(her beskrives processerne i detaljer, derfor betegnelsen UDDYBET VERSION, men et forsimplet indlæg under mappen 'juni' kan ligeledes læses med betegnelsen SIMPEL VERSION)
Fotosyntesen overordnet
Fotosynteseprocessen er noget ganske særligt, som kun planter har patent på. Det er den ældste og vigtigste biokemiske proces på jorden. Under processen omdannes de uorganiske stoffer vand (H2O) og kuldioxid (CO2) til det organiske stof glukose (C6H12O6), samt ilt (O2) og vand via solenergi (dvs. elektromagnetisk stråling). Overordnet kan processen beskrives således:
12 H2O + 6 CO2 + lysenergi → C6H12O6 + 6 O2 + 6 H20
Fotosyntesen er en såkaldt redoxproces (består af en reduktion, dvs. elektronoptagelse, og oxidation, dvs. elektronafgivelse). Dvs. at planten vha. lysenergien flytter elektroner fra oxygenatomerne i vandmolekylet til kulstofatomerne i CO2. Vand oxideres og CO2 reduceres. Når enzymerne flytter elektronerne, følger der protoner (H+-ioner) med. Det er strømmen af e- og H+ der driver de øvrige processer i fotosyntesen.
I planten foregår fotosynteseprocessen, der estår af adskillige biokemiske underprocesser, i plantecellernes grønkorn (også kaldet kloroplastre) inde i cellerne.
Planter optager kuldioxid fra luften via spalteåbninger på undersiden af blade i dagslys. Den dannede ilt udskilles ligeledes via spalteåbningerne. Vandet optages fra jorden via rødderne og den elektromagnetiske stråling optages direkte i farvestoffetstoffet klorofyl i plantecellerne. Da klorofyl er grønt, tilbagekastes en betydelig del af netop det grønne lys, hvilket giver bladene deres karakteristiske grønne farve.
Fotosynteseprocessen kan ydermere opdeles i primærprocessen eller lysprocessen, en lys-afhængig delproces, som finder sted i tylakoiderne i grønkornene i plantecellerne, samt sekundærprocessen, mørkeprocessen eller Calvin cyklus, en energikrævende, lys-uafhængige proces i, som foregår i plantecellernes stroma eller cytoplasma. Mørkeprocessen foregår i modsætning til lysprocessen uden direkte brug af lysenergi, hvilket har givet processen sit karakteristiske navn, og foregår uanset og det er lyst eller mørkt.
Ved lysprocessen absorberer kloroplastrenes farvestoffer, fotosyntesepigmenterne klorofyl a og b,
lysenergien, og oxiderer vand til O2, mens elektroner og protoner afgives. Samtidig leverer lysprocesserne energi i form af ATP til mørkeprocessen. I de efterfølgende mørkeproces reduceres CO2 til glukose vha. H+ og e-.
Lysprocessen og mørkeprocessen er koblede processer hvor lysprocessen leverer energi til syntesen af glukose i mørkeprocessen, og mørkeprocessen kan således kun forløbe samtidig med lysprocessen.
Kloroplasternes/grønkornenes opbygning
Kloroplastre er omgivet af to lag enhedsmembran. Det indra af kloroplasten er inddelt i to rum. Inden for membranerne befinder stroma sig, som er en cytoplasmalignende væske. I stroma ligger et kompliceret opbygget hulrum, lumen, som er omgivet af en enhedsmembran, thylakoidmembranen. En stor del af thylakoidmembranen er organiseret i grana der er stakke af flade sække.
I stromaet befinder de enzymer sig som udfører mørkeprocesserne.
I thylakoidmembranerne sidder tre typer af proteinkomplekser:
- Fotosystem I og Fotosystem II, som indeholder klorofyl og indfanger lyset.
- Et enzymkompleks som indeholder cytokrom bf og som transporterer proteoner, H+, igennem membranen.
- ATP-syntase, et enzym der indeholder en H+-kanal. Når der diffunderer H+ igennem den, danner enzymet ATP ud fra ADP + P.
Disse proteinkomplekser udfører tilsammen lysprocessen.
Fotosystemernes virkemåde i korte træk
Klorofyl og andre hjælpepigmenter befinder sig i fotosystemerne i thylakoidmembranen. Hvert fotosystem består af et lyshøstningskompleks og et reaktionscenter. Lyshøstningskomplekset er et proteinkompleks hvortil der er bundet et stor antal klorofylmolekyler, fotosystemets antennemolekyler. Centralt i fotosystemets reaktonscenter sidder to klorofylmolekyler.
Når antennemolekylerne modtager lysenergi af en passende bølgelængde, energiberiges en elektron i et af molekylerne (den eksiteres), hvilket bevirker, at elektronen herved kan overføres til et nabomolekyle. På den måde overføres der energi fra antennemolekyle til antennemolekyle indtil energien aktiverer klorofylmolekylerne i fotosystemets reaktionscenter. Det afgiver en elektron til en elektronacceptor som sender det videre.
Lysprocessen i detaljer
Fotosynteseprocessen starter i fotosystem II, når antennemolekylerne rammes af lys, således at reaktionscenteret aktiveres, som via cytokrom bf afgiver elektronerne til fotosystem I.
Lyset trækker på denne måde elektroner ud af fotosystem II. Elektronerne ertattes ved at enzymkompleks i forbindelse med fotosystem II oxiderer vandmolekyler, sender elektronerne til reaktionscenteret og afgiver H+ til lumen. Resten af vandmolekylet afgives som ilt, dvs. den dannede O2 under fotosyntesen er et restprodukt i processen.
Når cytokrom bf optager og afgiver elektroner, pumper det samtidig H+ fra stroma ind i lumen. Pumpen drives altså af elektronstrømmen. H+ kan diffundere tilbage igen gennem H+-kanalerne i ATP-syntasemolekylerne. Hver gang der diffunderer ca. 4 H+ gennem enzymet, dannes et ATP-molekyle. Man siger at ADP fosforyleres til ATP. Fordi fosforyleringen drives af lyset, kaldes den en fotoforsforylering.
ATP dannes i stromaet hvor dan leverer energi til dannelsen af glukose i mørkeprocesserne.
I fotosystem II optager antennemolekyler lysenergi, som siden videregives til det specifikke klorofylmolekyle P680, der afgiver en elektron ved optagelsen af en foton. Elektronen videregives herefter til en omdannet form af klorofyl, feofytin (mangler magnesiumatom, men har i stedet to brintatomer), der videregiver elektronen til et plastoquinonmolekyle. Denne reaktion bevirker, at der dannes en elektronstrøm langs en transportkæde. P680 skal bringes tilbage til sin stabile tilstand (det mangler jo en elektron), og derfor optager den en elektron fra elektrondonoren tyrosin Z, som frigiver et brintatom, hvorfra den nye elektron til P680 således stammer. Tyrosin Z genetablerer sin stabile tilstand ved at optage et brintatom fra et vandmolekyler (som kommer fra tylakoidets indvendige vægge). Tilbage fra vandmolekylet vil således være to iltatomer, dvs. O2 (ilt), hvilket, som nævnt, er årsagen til iltdannelsen under fotosyntesen. Plastoquinon frigiver herefter sine protoner til tylakoidets indre og afgiver sine elektroner til cytokromkompleks bf. De afgivne protoner i tylakoidets indre harmonerer ikke med antallet af protoner i stroma, så for at skabe en koncentrationsudligning, søger protonerne ud igennem en kanal skabt af ATP-syntaseenzymet, hvilket samlet danner ATP, der skal bruges til mørkeprocessen. Der dannes en såkaldt protongradient.
Samtidig slutter elektronstrømmen i transportkæden med at coenzymet NADP+ reduceres til NADPH via to elektroner og en H+-ion.
Fotosystem I virker i princippet som fotosystem II. Antennemolekylerne indfanger solenergi og overfører energien til reaktionscenteret, som afgiver elektroner, som denne gang erstattes med elektronerne fra fotosystem II. Elektronerne afgives videre til NADP+ som også binder H+ fra stroma og omdannes til NADPH, som tidligere nævnt. NADPH overfører H+ og e- til mørkeprocesserne. Fotosystem I er således ansvarlig for at afgive NADPH til stroma. I modsætning til fotosystem II, vender elektronerne ikke tilbage til det samme molekyle, så denne del af fotosynteseprocessen betegnes også som den ikke-cykliske fosforylering.
Fotosystem I starter ved, at en elektron fra cytokromkompleks bf optages af plastcyanin, som side
n videregives til klorofyl P700 (der opfanger fotoner ved bølgelængden 700nm). På samme måde som i fotosystem II opfanger antennemolekylerne energi, der videreføres til P700 som elektroner. P700 mister således en elektron, som dog erstattes af en elektron fra reaktionscenteret. Elektronen afgivet fra P700 optages af et nyt klorofylmolekyle, som igen videregiver elektronen til et nyt molekyle, ect. indtil elektronen når molekylet ferredoxin, bestående af to jernatomer og to svovlatomer, som forekommer tæt ved stroma mellem cytokromkomplekserne og fotosystem I. Molekylet er essentielt, idet elektronen herfra kan videregives til mange forskellige processer, men hvis vi antager, at det er fotosynteseprocessen vi lige nu har med at gøre, vil ferredoxin afgive elektronen til enzymet ferredoxin-NADP-reduktase i stroma. Enzymet binder to H+-ioner fra stroma samt NADP via elektronen fra ferredoxin.
I alt kan lysproccen skrives som:
12 H20 + 12 NADP+ + 48 H+ (i stroma) + 12 H+ (i stroma) → 6 O2 + 12 NADPH + 72 H+ (i lumen)
Når de 72 H+ diffunderer ud igen gennem thylakoidmembranen, danner ATP-syntase ca. 18 ATP:
18 ADP + 18 P + 72 H+ (i lumen) → 18 ATP + 72 H+ (i stroma)
Ud fra oxidationen af de 12 vandmolekyler der indgår i fotosyntesens reaktionsligning dannes der således 6 O2, 18 ATP og 12 NADPH + 12 H+. Men dette er kun lysprocessen. Efterfølgende anvendes den dannede energi ATP og NADPH i mørkeprocessen, den sidste del af fotosyntesen.
Undersøgelser af planten 'gåsemad' tyder desuden på, at planter kan tilpasse sig ændringer i lysets farvesammensætning ved at flytte proteinkomplekset LHC mellem fotosystem I og II, dvs. state transition. Læs dyberegående om dette
her.
Mørkeprocessen i detaljer (med udgangspunkt i C3-planter)
Mørkeprocessen består af en række biokemiske proesser som tilsammen udgør en cyklisk proces, Calvins cyklus, der kan inddeles i tre trin.
- CO2 som planten optager fra luften, bindes af ribulose-1,5-bifosfat som er et monosakkarid opbygget af 5 C-atomer. Molekylet spaltes ved processen til to molekyler glycerat-3-fosfat som hver indeholder 3 C-atomer via enzymet RuBisCO.
- NADPH afleverer H+ og e- hvorved glycerat-3-fosfat reduceres til glyceroladehyld-3-fosfat (glucid). Denne proces kræver ud over NADPH også ATP. 2 glycerolaldehold-3-fosfat kan omdannes til triosefosfat (G3P) der videre kan omdannes til glukose, som endvidere kan omformes til andre kulhydrater samt aminosyrer og lipider, som skal bruges af planten (bl.a. til oplagring i form af stivelse eller til respirationsprocessen). Fra Calvin cyklus kan dog også afgives fructose-6-fosfat i stedet for G3P, ved at to molekyler glycerolaldehyld-3-fosfat sammenkobles, som omdannes til glukose og siden stivelse i grønkornene.
- Størstedelen af glycerolaldehyld-3-fosfat-molekylerne bruges imidlertid til at gendanne ribulose-1,5-bifosfat, så der kan bindes ny CO2 via en række processer, der kræver ATP.
Fotosyntese og livsvilkår hos planter
Fotosyntesens størrelse vil være afhængig af lysmængden, mængden af CO2 og H2O og hvor mange fotosyntesepigmenter og -enzymer planten indeholder.
Planter som er tilpasset livet på skyggefulde steder, fx i skovbunden, indeholder alt i alt færre fotosyntesepigmenter og -enzymer end planter tilpasset solbeskinnede levesteder. Det er især mængden af klorofyl og RuBisCO som er nedsat, og deres maksimale fotosyntesehastighed er derfor tilsvarende lavere. Til gengæld er der relativt flere pigmentmolekyler i forhold til de øvrige proteiner i kloroplastrene. Derfor vil de have en højere netoofotosyntese ved lavere lysintensiteter end lysplanterne. Ofte indeholder skyggeplanter desuden flere hjælpepigmenter, ud over klorofyl a og b, som sætter dem i stand til at absorbere lys ved bølgelængder som passerer de øvrige planters blade.
Lysblade har en større respiration end skyggeblade. Derfor vil deres nettofotosyntese være mindre end skyggebladenes ved ganske lave lysintensiteter. Nettofotosyntesen vil være negativ hvis lysintensiteten har en værdi hvor fotosyntesen bliver mindre end respirationen. En lysintensitet, hvor fotosyntesen og respirationen er lige store, er plantens såkaldte lyskompensationspunkt.
Respiration hos planter - overordnet
Ved fotosyntesen binder planterne energi i kulhydraternes kemiske bindinger. Energien kan frigøres igen ved forbrænding i planten selv, nemlig via respirationen - en direkte modsætning til fotosyntesen i planterne. Det er nødvendigt for planter at optage ilt for at kunne gøre den dannede energi fra fotosyntesen tilgængelig i de dele af planten, hvor der ikke forekommer fotosyntese, f.eks. i spirende frø og om natten. Respirationsprocessen hos planter finder således sted både om dagen og om natten, mens fotosyntesen kun forekommer om dagen. Processen finder sted i planternes mitokondrier i plantecellerne, og langt de fleste levende væsner anvender respiration for at udvinde bioenergi af organisk stof. Ved respirationen nedbrydes glukose derfor ved hjælp af ilt, så den kemiske energi i glukose overføres til ATP. Ved processen dannes desuden kuldioxid og vand. Processens kemiske reaktionsligning ser således ud:
C6H12O2 + 6 O2 → 6 CO2 + 6 H2O + ATP (energi)
Overordnet set nedbrydes glukosen fra fotosyntesen til CO2 og H+ + e-. H+ (protoner) og e- (elektroner) overføres til O2 hvorved der dannes H2=. Dvs., at C oxideres (afgiver e-) mens O reduceres (optager e-). Den energi der frigøres ved respirationsprocessen, bruges til at danne ADP og P til ATP, som overfører den frigjorte energi til andre stoffer i cellen.
Respirationsprocessen i dybden
Respirationsprocessen kan inddeles i tre hovedprocesser: glykolysen, citronsyrecyklussen og elektrontransportkæden.
I glykolysen og citronsyrecyklussen nedbrydes glukose. H+ og e- overføres til O2 i elektrontransportkæden og der dannes H2O. Glykolysen foregår i cellens cytoplasma, mens citronsyrecyklussen og elektrontransportkæden foregår i cellens mitokondrier. Citronsyrecyklussen benævnes desuden også Krebs cyklus efter Hans Krebs (første person som beskrev processen) og elektronstransportkæden kendes også som respirationskæden.
Hvis cellen mangler ilt vil der ingen respiration forekomme, men en gæring i cellens cytoplasma vil foregå istedet. Ved en gæring vil enzymer i cytoplasma danne laktat efter glykolysen, og udbyttet i form af ATP bliver lavere.
Glykolysen

Processen består af ni delprocesser. Ved de første to trin lades glukosen op ved at der overføres 2 P fra ATP til molekylet, dvs. glukosen fosforyleres. Herefter spaltes glukosen i to C-3-forbindelser der hver fosforyleres med yderligere en P. I det efterfølgende trin overføres P til ADP hvorved der dannes fire ATP-molekyler. Nettoudbyttet af glykolysen af derved 2ATP.
I glykolysen fraspaltes endvidere 4 H+ og 4 e- fra glukosen. De optages dernæst af coenzymet NAD+ som overfører dem til mitokondrierne til den videre citronsyrecyklus og elektrontransportkæde.
Under glykolysen omdannes glukosen således til to molekyler pyrovat i cytosolen, som er et helt centralt molekyle i de efterfølgende processer, da de transporteres videre ind i mitondrierne til oxidation.
Sammenfattende kan glykolysen beskrives som:
C6H12O6 + 2 NAD+ + 2 ADP + 2 P -> 2 CH3COCOO- (pyrovat) + 2 NADH + 2 H+ + 2 ATP
Citronsyrecyklussen og mitokondriernes opbygning
For at forstå først citronsyrecyklussen, og siden elektrontransportkæden, er mitrokondriernes opbygning essentiel af forstå. Ligesom kloroplastre i planten er mitokondrierne opbygget af to lag enhedsmembran. Enzymerne som katalyserer processerne i citronsyrecyklussen befinder sig i mitokondriernes såkaldte matrix, hvilket er et område der befinder sig inden for den inderste foldede membran. Elektrontransportkæden består af en række enzymkomplekser der sidder i mitokondriernes indre membran. Processerne foregår altså i samme celleorganel, men forskellige steder heri.
Hos prokaryoter vil de to processer dog foregår i cytoplasmaet hos cellen.
Er der ilt til stede vil pyrovatet (dannet under glykolysen) blive transporteret ind i mitokondrierne hvor resten af respirationsprocessen foregår. Der er således tale om en aerob oxidation. Dette sker ved, at pyrovat bindes til Coenzym A, forkortet CoA eller HSCoA. Ved bindingen til CoA spaltes CO2 og 2e- fra hver pyrovat som omdannes til en acetylgruppe, så resultatet er dannelse af stoffet acetyl-CoA, der indgår i citronsyrecyklussen. Acetyl-CoA overfører hernæst acetylgruppen til stoffet oxaloacetat der herved omdannes til citat. Ved processen spaltes et molekyle H2O for hvert acetyl-CoA. De 2 e- og 2 H+ fra vandmolekylerne overføres af CoA til NAD+ som videreoverfører dem til elektrontransportkæden.
Gennem otte delprocesser omdannes citat igen til oxaloacetat, og undervejs bindes der yderligere 4 H2O samt fraspaltes yderligere 4 CO2. Desuden afgives der i alt 12 H+ og 12e- til elektronacceptorerne NAD+ og FAD. FAD er ligesom NAD+ et coenzym der overfører H+ og e- mellem enzymer. Derved dannes FADH2 og NADH. Endelig dannes der også 2 ATP ved processerne.
Citronsyrecyklussen er en cyklisk proces, og som tidligere nævnt starter og slutter processen med dannelsen af oxaloacetat. Herfra kan oxaloacetat igen dehydrogeneres til citrat via aconitaseenzymer, så der dannes vand som restprodukt. Citratet omdan

nes til cis-Aconitat via en hydrogenering hvor samme enzymgruppe og vand indgår i reaktionen. Herefter fortsætter omdannelsen af cis-Aconitat til Isocitat via en oxidation af cis-Aconitat via isocitatdehydrogenaseenzymer, hvor coenzymet er NAD+ og der foruden isocitat også dannes NADH+. Efterfølgende forekommer der en decarboxyering af isocitatet via samme enzymgruppe, så der dannes oxalosuccinat. Processen er dog på ingen måde fuldendt, og fortsætter med en omdannelse af oxalosuccinat til alfa-ketoglutarat. Denne delproces er forårsaget af alfa-ketoglutaratdehydrogenaseenzymer, med NAD+ og CoA-SH som coenzymer, og selve omdannelsen er en oxidativ decarboxylering. Ud over omdannelsen af oxalosuccinat til alfa-ketoglutarat dannes også NADH+. Alfa-ketoglutarat hydrolyseres hernæst via succinyl-CoA syntetaseenzymer, med GDP + P som coenzymer, så resultatet er dannelsen af succinyl-CoA samt GTP og CoA-SH. Succinyl-CoA oxideres via succinatdehydrogenaseenzymer, med FAD som coenzym, og herved omdannes succinyl-CoA til succinat (desuden dannes der også FADH2). Ved næste trin i cyklussen indgår der vand som coenzym, hvormed der forekommer en addition af succinat. Resultatet er dannelsen af fumarat via fumaraseenzymer. Næstsidste delproces i citronsyrecyklussen består af en oxidation af fumarat således, at der dannes L-Malat via malat dehydrogenaseenzymer (hvor NAD+ er coenzym). Desuden dannes der også her NADH og H+. Til sidst når vi tilbage til udgangspunktet: oxaloacetat. Dette dannes via en kondensation via citratsyntaseenzymer. Citronsyrecyklussen kan starte på ny.
Sammenfattet kan citronsyrecyklussen derfor skrives som:
2 CH3COCOO- + 2 H+ + 6 H2O + 8 NAD+ + 2 FAD + 2 ADP + P ->
6 CO2 + 8 NADH + 8 H+ + 2 FADH2 + 2 ATP
Det kan altså ud fra ovenstående reaktionsskema ses, at det ikke er ud fra citronsyrecyklussen, at ATP dannes (hvis man kigger samlet på respirationsprocessen). Tværtimod oxideres to carbonatomer i processen til CO2, som udåndes, og energien fra denne oxidation oplagres som GTP, NADH og FADH2. De to sidstnævnte er som bekendt coenzymer, og de anvendes derfor videre i den oxidative fosforylering, i elektrontransportkæden.
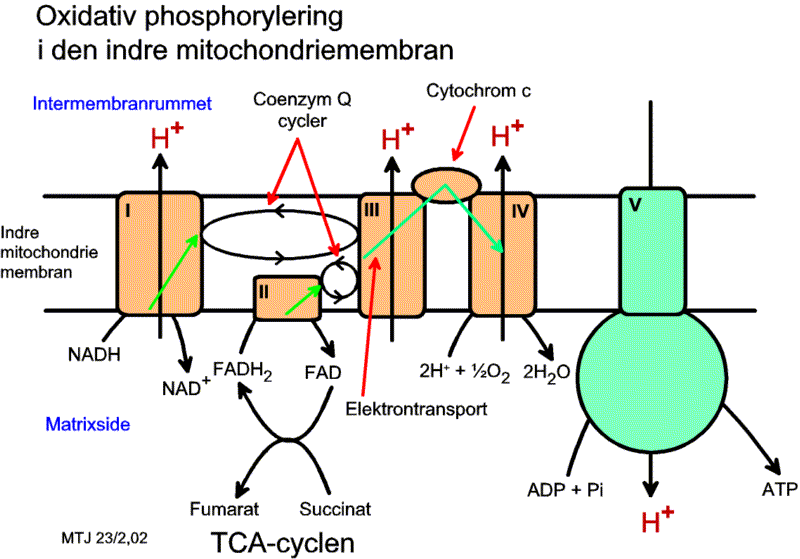
Elektrontransportkæden
Det er faktisk først i denne proces, at ilttilstedeværelsen bliver særlig aktuel. Kæden består som tidligere nævnt af en række enzymkomplekser der forekommer i mitokondriernes indre foldede membran. Enzymerne har den egenskab, at de kan binde og afgive elektroner, dvs. at de skiftevis kan reduceres og oxideres - dvs. en række redoxprocesser. De danner således en kæde af enzymer der transporterer elektroner, heraf navnet, og energien fra oxidationsprocesserne bruges til at pumpe H+ gennem membranen. Elektrontransportkæden består af fem store komplekser: enzymkompleks I, II, III, IV og ATP-syntase. Ud over disse makromolekyler forekommer også mindre molekyler som cytochromer og ubiquinon, der koblede transporterer elektronerne igennem kæden for i slutningen af overføre dem til O2, der dernæst reagerer med H+ hvorved der dannes vand. Flere af de fem store enzymkomplekser minder meget i opbygning og virkemåde som enzymkomplekserne i fotosyntesens lysprocesser.
Enzymkompleks I kan binde NADH+ og H+ samt et antal H+ fra mitokondriets matrix. Herefter kan komplekset overføre de 2 H+ + 2e- til enzymkompleks III. Energien der frigøres ved dette, anvender enzymkomplekset til at pumpe H+ gennem membranen fra matrix til intermembranrummet, dvs. væsken mellem mitronkriets indre og ydre membran. Enzymkompleks I pumper 4 H+ gennem membranen for hvert NADH den oxiderer.
Enzymkompleks II kan spalte 2H+ + 2e- fra FADH2. De 2 H+ + 2e- overføres ligeledes til enzymkompleks III, men her sker der ikke en udpumpning af H+.
Enzymkompleks III kan således optage H+ + e- fra enzymkompleks I og II. Enzymkompleks III pumper 2 H+ til intermembranrummet og afleverer 2e- til enzymkompleks IV.
Enzymkompleks IV optager elektroner fra enzymkompleks III. Disse elektroner overføres sammen med H+ fra mitokondriets matrix til O2 som derved omdannes til H2O. Den energi der frigøres ved processen, bruger enzymkompleks IV til at pumpe H+ fra matrix til intermembranrummet. Det pumper 4 H+ igennem for hvert 2e- det modtager.
Samlet set vandrer der derved elektroner fra NADH og FADH2 gennem enzymkomplekserne som skiftevis reduceres og oxideres. Til sidst optager O2 elektronerne og reduceres. Sammen med elektronerne bindes O2 til H+ og som resultat dannes H2O. Den energi der frigøres ved denne proces, driver udpumpningen af H+.
For hvert NADH + H+ der overføres til elektrontransportkæden, pumpes der 10 H+ gennem membranen, 4 fra enzymkompleks I, 2 fra enzymkompleks III og 4 fra enzymkompleks IV.
For hver FADH2 pumpes der 6 H+ igennem, 2 fra enzymkompleks III og 4 fra enzymkompleks IV.
Transporten af elektronerne igennem elektrontransportkæden skaber via de mange små enkeltreaktioner en samlet stor reaktion - en dannelse af en større mængde energi. Denne energi omsættes af enzymer til transport af H+ fra mitonkodriematrix til rummet mellem inder- og ydermembranen, hvilket opbygger en protongradient. Det er denne protongradient, som siden udnyttes til dannelse af det energiholdige molekyle ATP i respirationsprocessen. For at danne ATP skal det sidste enzymkompleks i spil, nemlig ATP-syntase. Enzymkomplekset indeholder en H+-kanal. Når H+ diffunderer gennem kanalen tilbage til matrix bruger enzymet energi en til netop at danne ATP. Enzymkomplekset omsætter altså så at sige protonernes opsparede kemiske energi til ATP når de føres igennem H+-kanalen i ATP-syntase. Der skal bruges ca. 4 H+ for at danne et ATP-molekyle og få det transporteret ud i cellens cytoplasma. For hvert NADH-molekyle der overføres til elektrontransportkæden, dannes der derfor ca. 2,5 ATP, mens der dannes ca. 1,5 ATP for hvert FADH2-molekyle. Dannelsen af ATP betegnes også den oxidative fosforylering.
Nettoligningen for respirationskæden er samlet:
6 O2 + 10 NADH + 10 H+ + 2 FADH2 + 28 ADP + 28 P ->
12 H2O + 10 NAD+ + 2 FAD + 28 ATP
To af NADH-molekylerne blev dog dannet ved glykolysen i cytoplasma. De optages i mitokondriet ved aktiv transport hvilket er omkosntingsfuldt for cellen - ca. 1 ATP for hvert molekyle NADH der skal transporteres.
En suveræn illustration til ovenforbeskrevne proces kan ses her:
For en endnu mere dybdegående beskrivelse af elektrontransportkæden kan du også se:
Følgende dokument giver desuden også en rigtig god beskrivelse af henholdsvis Calvin cyklus, glykolysen og citronsyrecyklussen (med tilhørende gode illustrationer):



{kind=link}
{kind=link}